大椎龍
| 大椎龍 |
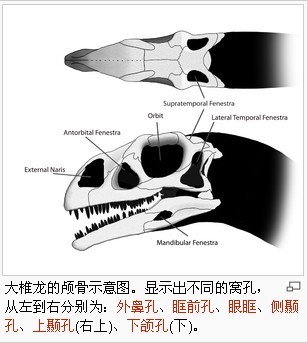 |
大椎龍(屬名:Massospondylus),又名巨椎龍,屬名在希臘文意為巨大的脊椎。大椎龍是原蜥腳下目的一屬,生存於早侏羅紀赫塘階到普林斯巴赫階,約2億年前到1億8300萬年前。大椎龍是在1854年,由理查德·歐文根據來自於南非的化石而命名。因此它們是最早命名的恐龍之一。大椎龍的化石已經在南非、萊索托、以及贊比亞等地發現。
目錄
簡介
大椎龍的頭部小,長度接近股骨長度的一半。頭部的眾多窩孔,減低了頭部的重量,並提供肌肉附着處,與容納感覺器官。這些窩孔在頭部兩側成對排列。頭部前方為兩個大型、橢圓形的鼻孔。眶前孔(Antorbital fenestrae)位於鼻孔與眼睛之間,小於板龍的眶前孔。眼窩所占頭部面積比例大於板龍的眼窩。頭部後方則為兩對顳顬孔(Temporal fenestrae):位於眼窩後方中間的側顳孔(Lateral temporal fenestrae)、與頭頂上側的上顳孔(Supratemporal fenestrae)。下頜兩側也有小型窩孔。傳統上,大椎龍的頭部被重建成較板龍的為寬、短,但從不同的標本得知,這個形狀可能是導因於化石被擠壓時,遭受到不同的力道而變形。有些頭部特徵隨者個體而有所不同,例如:眼窩上側的骨頭厚度、上頜骨後段的高度。這些差異可能是兩性異形,或者是個體變化。
評價
大椎龍的第一個化石是在1953年發現,由約瑟夫·奧賓(J. M. Orpen)發現於南非哈利史密斯的上艾略特組。並在1954年,由古生物學家理查·歐文(Richard Owen)命名,大椎龍的正模標本包含:從頸部到背部與尾巴的脊椎、一個肩帶、一個肱骨、部分骨盆、一個股骨、一個脛骨、以及手掌與腳掌的骨頭。這個正模標本存放於倫敦的英國皇家外科醫學院,但在第二次世界大戰期間遭到摧毀,只有部分還在。在南非與萊索托的上艾略特組、克萊倫斯組、布什維爾砂岩組,發現了大椎龍的可能化石;其他地區還有:津巴布韋上卡羅砂岩層的森林砂岩組、阿根廷的Cañon del Colorado組或El Tranquilo組、以及亞利桑那州的卡岩塔組。這些化石由至少80個部分骨骸、以及4個頭顱骨所構成,幼年與成年個體都有。
在1985年,亞利桑那州卡岩塔組發現一個頭顱骨,可能屬於大椎龍。這個卡岩塔頭顱骨與非洲所發現的最大型頭顱骨相比,還大上25%。前上頜骨具有4顆牙齒,上頜骨有6顆牙齒。齶骨有長約1公厘的迷你牙齒,這在恐龍中相當獨特。在亞利桑那州卡岩塔組發現的頭顱骨,比南非發現的最大頭顱骨,還大上25%。對於非洲大椎龍頭顱骨的重新研究,指出卡岩塔組的標本並不屬於大椎龍。
阿根廷過去曾發現疑似大椎龍的化石,包含數個部分骨骸,與至少一個頭顱骨,發現於聖胡安省的Cañon del Colorado組,年代為侏羅紀早期。在2009年,這些化石被認為屬於大椎龍的近親,被建立為獨立屬,遠食龍(Adeopapposaurus)。模式種是刀背大椎龍(M. carinatus),在過去150年來,還有其他七個種被建立過,其中只有M. kallae是有效種。在過去幾年,原蜥腳下目的系統分類學已被多次更改過,而許多科學家對於大椎龍在恐龍演化樹中的正確位置,仍沒有一致的結論。大椎龍科被建立以包含大椎龍屬,但由於原蜥腳類恐龍之間的關係沒有定論,所以無法確定有哪些恐龍屬於大椎龍科。在2007年,有數個研究支持科的有效性。
種屬
大椎龍的模式種是刀背大椎龍(M. carinatus),是由理查·歐文所建立。屬名衍化自古希臘文,masson/μάσσων意為"巨大的",而sphondylos/σφονδυλος意為"脊椎";而種名意為"剃刀般的背部" 。在2009年,保羅·巴雷特(Paul Barrett)將一個發現於南非上艾略特組的部分頭顱骨,命名為新種M. kaalae。上艾略特組也發現過一些刀背大椎龍的化石,顯示這兩個種生存於相同年代。這兩個種的頭顱骨形態、特徵不同,例如M. kaalae的前上頜骨比例較長。
大椎龍過去曾有過許多其他種,但大部分不被承認是有效種,包括:
布朗氏大椎龍(M. browni):由哈利·絲萊(H.G. Seeley)在1895年命名。
哈里斯氏大椎龍(M. harriesi):由羅伯特·布魯姆(Robert Broom)在1911年命名。
希氏大椎龍(M. hislopi):由理查德·萊德克(Richard Lydekker)在1890年命名.
休尼氏大椎龍(M. huenei): 由庫柏(M. R. Crooper)在1981年命名.
拉氏大椎龍(M. rawesi):由萊德克在1890年命名。
施氏大椎龍(M. schwarzi): 由席尼·賀頓(Sydney H. Haughton)在1924年命名。
布朗氏大椎龍、哈里斯氏大椎龍、以及施氏大椎龍,都發現於南非角省的上艾略特組。三者都只有破碎的化石,大部分近年的研究將它們視為疑名。布朗氏大椎龍的化石有2節頸椎、2節背椎、3節尾椎、以及混雜的後肢碎片。哈里斯氏大椎龍的化石只有前肢。施氏大椎龍的化石是一個不完整後肢與薦骨。希氏大椎龍與拉氏大椎龍的化石都發現於印度。希氏大椎龍的化石為脊椎,發現於安得拉邦的馬勒爾組,年代為晚三疊紀。拉氏大椎龍的化石是兩顆牙齒,發現於馬哈拉施特拉邦的Takli組,年代為晚白堊紀。在最近的看法裡,希氏大椎龍暫時被視為一種無法確定的蜥腳形亞目動物。拉氏大椎龍可能是種獸腳類恐龍,或者並非恐龍。庫柏則提出休尼氏大椎龍就是許氏祿豐龍,因此他認為祿豐龍與大椎龍是同種動物。但這個看法並不被接受。
疑名
有數種恐龍常被認為是大椎龍的異名,包括:Aristosaurus、Dromicosaurus、Gryponyx、Hortalotarsus、Leptospondylus、以及Pachyspondylus,這些恐龍的有效性不大,也被視為疑名。Hortalotarsus是由哈利·絲萊在1894年建立,化石包含部分腿骨。第二年,理察·歐文將一些脊椎命名為Leptospondylus與Pachyspondylus。這些化石都在第二次世界大戰期間遭到摧毀。在1920年,E.C.N. van Hoepen根據一個接近完整的骨骸,建立了Aristosaurus。他另外將一個部分骨骸命名為Dromicosaurus。在1924年,席尼·賀頓命名了Gryponyx,化石為臀部骨頭。以上化石都發現於南非的赫塘階到錫內穆階,約跟大椎龍同一時期。根據國際動物命名法規,這些名稱是大椎龍的次同物異名。大椎龍比這些名稱還早出現在科學文獻中,因此大椎龍具有優先權。
分類
大椎龍屬於原蜥腳下目,原蜥腳下目是群早期蜥臀目恐龍,生存於三疊紀與侏羅紀,並在侏羅紀末期滅亡。原蜥腳下目的其他著名屬包含:板龍、雲南龍、與里奧哈龍。基礎蜥腳形亞目的系統發生學仍在爭論中,根據親緣分支分類法,一個天然演化支應包含它們的共同祖先與其共同祖先的所有後代,所以許多過去被認為是典型原蜥腳類的物種,因為無法與原有物種構成一天然演化支,近年被排除在原蜥腳下目之外。但哪些物種構成原蜥腳類為一個單系群,仍不確定。在2003年,亞當·耶茨(Adam Yates)與詹姆斯·基欽(James Kitching)公布了一個演化支,包含:里奧哈龍、板龍、科羅拉多斯龍、大椎龍、以及祿豐龍。在2004年,彼得·加爾東(Peter M.Galton)與保羅·阿普徹奇(Paul Upchurch)則將原蜥腳下目列為單系群,包含:砂龍、近蜥龍、愛珍多龍、卡米洛特龍、科羅拉多斯龍、優肢龍、金山龍、萊森龍、祿豐龍、大椎龍、黑丘龍、鼠龍、板龍、里奧哈龍、呂勒龍、農神龍、鞍龍、槽齒龍、易門龍、以及雲南龍。在2005年,傑佛瑞·威爾森(Jeffrey A. Wilson)提出大椎龍、金山龍、板龍、與祿豐龍,形成一個天然演化支,可能還有貝里肯龍、雷前龍等蜥腳類恐龍。在2007年,Matthew F. Bonnan與耶茨提出卡米洛特龍、貝里肯龍、黑山龍可能屬於蜥腳下目。同樣在2007年,耶茨將雷前龍、黑丘龍、貝里肯龍歸類為基礎蜥腳類恐龍,而且認為原蜥腳下目是板龍科的異名,而沒有使用這分類。但耶茨並沒有排除少部分原蜥腳類構成一個單系群的可能性,這些成員包含板龍、里奧哈龍、大椎龍、以及它們的最近親。
大椎龍是大椎龍科的模式屬,該科名稱即來自於大椎龍。大椎龍科可能包含雲南龍,但在2007年,Lu等人將雲南龍列入個別的雲南龍科。在2007年,耶茨提出近蜥龍類,而大椎龍科的大椎龍、科羅拉多斯龍、祿豐龍,以及雲南龍屬於近蜥龍類。同樣在2007年,納森·史密斯(Nathan D. Smith)與迪亞戈·玻爾(Diego Pol)在他們的系統發生學研究中,將大椎龍、科羅拉多斯龍、祿豐龍、以及他們新建的冰河龍(Glacialisaurus),列入大椎龍科中。數個發現於阿根廷的疑似大椎龍化石,被建立為新屬,遠食龍(Adeopapposaurus),屬於大椎龍科。
古生態學
在早侏羅紀時期,世界各地的動物群與植物群都很類似。蕨類適應炎熱的氣候,成為常見的植物,而各地的恐龍動物群主要由原蜥腳類與基礎獸腳類恐龍組成。早侏羅紀時期的非洲南部應該是沙漠環境。與大椎龍共存於非洲的同時期動物包含:早期的鱷形超目動物、獸孔目的三瘤齒獸科與三棱齒獸科、哺乳類的摩爾根獸科,其他的恐龍則包含:小型獸腳類合踝龍、黑丘龍的一種、早期的鳥臀目萊索托龍、畸齒龍科的醒龍、畸齒龍、與狼鼻龍 。
目前不清楚哪些掠食性動物會以大椎龍為食。目前在非洲南部發現的早侏羅紀獸腳類恐龍,例如合踝龍,大部分比大椎龍還小。經推測,這些小型掠食動物使用快速攻擊的方式,消耗原蜥腳類恐龍的體力,而原蜥腳類恐龍會以手與腳上的大型指爪,來防衛自己。在2006年,南非艾略特組發現了一種新的獸腳類恐龍,龍獵龍(Dracovenator),身長約6米,生存於赫塘階到錫內穆階,約跟大椎龍同一時期。
生活習性
如同所有恐龍,大椎龍的許多生物學層面仍然未知,例如:行為、外表顏色、生理機能。但是近年的研究提出了關於生長模式、食性、步態、繁衍、以及呼吸等方面的假設。
一個2007年的研究提出大椎龍會將短前肢用在抵抗掠食動物、物種內打鬥、或是協助進食,但它們的前肢太短,無法碰到嘴部。科學家們假設大椎龍會使用拇指指爪來打鬥,或是折下樹上的食物。
生長模式
一個2005年的研究指出,大椎龍的姐妹分類單元-板龍的生長模式受到環境的影響。這個研究當食物為植物,或是氣候適宜時,板龍的生長速率會加快。這種生長模式稱為"發育可塑性"(Developmental plasticity)。但包含板龍的近親大椎龍在內的其他恐龍,都沒發現過這種現象。這個研究認為大椎龍有者特殊的生長模式,它們的生長率有少許變化,並依個體的體型而定。另一個研究則認為大椎龍的成長率可達增加每年34.6公斤,並持續約15年。。
食性
大椎龍屬於原蜥腳下目,原蜥腳類恐龍可能是草食性或雜食性動物。在80年代,科學家們開始爭論原蜥腳類是肉食性的可能性。但是,原蜥腳類是肉食性的假說已被否定,所有的近年研究傾向於原蜥腳類是草食性或雜食性。在2004年,加爾東與阿普徹奇發現大部分原蜥腳類的頭部特徵(例如頜部關節),較類似草食性爬蟲類,而非肉食性爬蟲類;牙齒形狀類似現代鬣蜥,而鬣蜥為草食性或雜食性動物。它們的齒冠最寬處大於齒根寬度,形成切割用邊緣,類似現代草食性或雜食性爬蟲類的牙齒。在2000年,保羅·巴雷特(Paul M. Barrett)提出原蜥腳類除了以植物為食外,還會以小型動物或屍體補充食物。曾經在南非的大椎龍化石附近發現胃石,維吉尼亞州的三疊紀晚期地層曾一個類似大椎龍的化石,也發現胃石。顯示大椎龍吞下石頭以協助消化。
繁衍
在1977年,詹姆斯·基欽(James Kitching)在南非金門高地國家公園發現了7顆1億9000萬年前的蛋化石,基欽鑑定它們極可能屬於大椎龍。直到將近30年之後,才能將蛋化石中的15厘米大胚胎取出。它們是目前所發現最古老的恐龍胚胎。這些接近出生的胚胎沒有牙齒,顯示它們沒有辦法自己覓食。根據這些胚胎缺乏牙齒,以及身體的比例,科學家們推測大椎龍出生後必定會有親代養育。這些接近出生的胚胎,四肢長度一樣,顯示它們剛孵化時是採取四足方式行走。與身體相比,頭部與眼睛很大,這點與其他脊椎動物的年幼個體相同。這些胚胎的四足性,顯示之後的蜥腳類恐龍會採取四足步態,是因為成年個體保有了幼年個體的特徵,這種演化現象名為幼體性熟(Pedomorphosis)。
呼吸系統
許多蜥臀目恐龍的脊椎與肋骨具有空腔,可以減輕骨頭的重量,或可能是初步的空氣流通系統,類似現代的鳥類。在這個系統中,頸部的脊椎與肋骨內有頸部氣囊,背椎前段內有肺臟,背椎後段與薦椎則有腹部的氣囊。這些器官運作者複雜、非常有效的呼吸作用。原蜥腳下目是蜥臀目中,唯一沒有額外氣孔的主要生物群。雖然板龍與槽齒龍的化石已發現可能的氣洞,這些凹洞非常的小。在2007年,一個研究提出原蜥腳類恐龍,可能如同它們的姐妹分類單元(獸腳亞目與蜥腳下目),具有頸部與腹部氣囊。這個研究認為很難確認原蜥腳類的骨頭是否有類似鳥類的肺臟,但可確定它們具有氣囊。[1]