纖維細胞
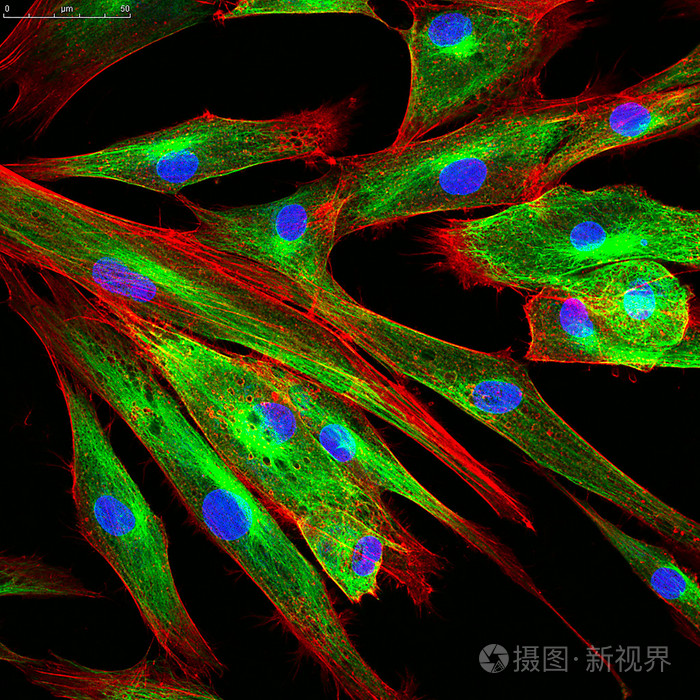 |
纖維細胞(fibrocyte)是機能不活躍的成纖維細胞。胞體呈梭形,胞質較少,弱嗜酸性,胞核小,染色深。電鏡下可見,細胞中粗面內質網和高爾基體均不發達。當組織受損時,纖維細胞可轉化為成纖維細胞而參與修復過程。[1]
目錄
簡介
纖維細胞和成纖維細胞是處於不同功能狀態的同一種細胞,如在組織損傷後的修復過程中,纖維細胞可轉化為功能活躍的成纖維細胞。纖維細胞較小,呈多角形,胞質較少,弱嗜酸性,胞核亦較小,染色較深,電鏡下粗面內質網較少,高爾基複合體不發達。成纖維細胞較大,呈星形或梭形,有突起,細胞核呈卵圓形,染色質稀疏,染色淡,核仁明顯,胞質較多。成纖維細胞能合成和分泌膠原蛋白、彈性蛋白和蛋白多糖,形成膠原纖維、彈性纖維和網狀纖維以及基質成分。[2]
轉變
在結締組織中,成纖維細胞還以其成熟狀態—纖維細胞(fibrocyte)的形式存在,二者在一定條件下可以互相轉變。 不同類型的結締組織含成纖維細胞的數量不同。通常,疏鬆結締組織中成纖維細胞的數量比同樣體積的緻密結締組織中所含成纖維細胞的數量要少,故分離培養成纖維細胞多以真皮等緻密結締組織為取材部位。
形態
成纖維細胞形態多樣,常見的有梭形、大多角形和扁平星形等,其形態尚可依細胞的功能變化及其附着處的物理性狀不同而發生改變。成纖維細胞胞體較大,胞質弱嗜鹼性,胞核較大呈橢圓形,染色質疏鬆着色淺,核仁明顯。電鏡下,其胞質可見豐富的粗面內質網、游離核糖體和發達的高爾基複合體,表明它具有合成和分泌蛋白質的功能。成纖維細胞尚可合成和分泌膠原纖維、彈性纖維、網狀纖維及有機基質。它合成的前膠原蛋白分子經內切酶作用,聚合和重排,可形成與成骨細胞合成分泌的膠原原纖維一樣具有64nm(640?)周期橫紋的膠原原纖維,膠原原纖維經互相粘合形成膠原纖維。經檢測,這兩種細胞合成分泌的膠原纖維均是Ⅰ型膠原纖維,在形態和生化結構上完全相同。 處於成熟期或稱靜止狀態的成纖維細胞,胞體變小,呈長梭形,粗面內質網和高爾基複合體均不發達,被稱為纖維細胞。在外傷等因素刺激下,部分纖維細胞可重新轉變為幼稚的成纖維細胞,其功能活動也得以恢復,參與組織損傷後的修復。另外,在結締組織中,仍保留着少量具有分化潛能的間充質細胞,它們在創傷修復等情況下可增殖分化為成纖維細胞。
作用
創傷修復
創傷均會造成不同程度的細胞變性、壞死和組織缺損,必須通過細胞增生和細胞間基質的形成來進行組織修復。在此修復過程中,成纖維細胞起着十分重要的作用。以傷口癒合過程為例,成纖維細胞通過有絲分裂大量增殖,並從4~5天或6天開始合成和分泌大量的膠原纖維和基質成分,與新生毛細血管等共同形成肉芽組織,填補傷口組織缺損,為表皮細胞的覆蓋創造條件。在傷口癒合中,成纖維細胞主要來源於真皮乳頭層的局部成纖維細胞和未分化的間充質細胞,以及血管周圍的成纖維細胞和周細胞內臟損傷時,參與修復過程的成纖維細胞多來自間質和包膜,以及粘膜下或漿膜下層的結締組織。有人認為創傷癒合過程中傷處聚集的大量成纖維細胞,一方面是由成纖維細胞通過分裂增殖而來,另一方面,更多地是由鄰近的間充質細胞、纖維細胞和毛細血管周細胞等演變或遊走到傷處。在創傷修復的後期,成纖維細胞通過分泌膠原酶參與修復後組織的改建。在某些病理條件下,以成纖維細胞為主要細胞成分的肉芽組織或增生組織塊還可以在非骨組織內發生鈣化,引起異位骨化( ectopicossification)。但對於異位骨化的參與細胞及其機制尚不十分清楚,未分化間充質細胞、成纖維細胞、內皮細胞和毛細血管周細胞等可劃歸為誘導性骨祖細胞的細胞都有可能參與這一過程。
骨創傷修復
最簡單和常見的骨創傷即是骨折,其癒合過程須經過炎性反應、清掃、纖維骨痂和骨性骨痂4個階段。不同階段參與的細胞主體不同。成纖維細胞從骨折第3天起就出現於骨折局部血腫中,骨折後5天即在機化血腫及骨折斷端的間隙及其周圍大量存在,是參與纖維骨痂階段的主要細胞成分。在此階段成纖維細胞一方面大量分裂增殖,一方面又合成和分泌大量I型膠原,使肉芽組織逐步變成疏鬆的結締組織,將骨斷端包圍起來,形成接合兩骨折斷端的巨大的纖維骨痂然而,這種由無數成纖維細胞和豐富的肉芽組織為主體構成的纖維結締組織卻不會演變為在其他組織創傷修復時常見的癜痕組織,而是通過鈣鹽結晶在其內部不斷沉積,逐漸演變為骨性骨痂,使骨折局部的修復達到骨性癒合,恢復骨組織的結構。此時,骨折癒合部只有骨組織而不再存在成纖維細胞。
成骨作用
對骨折局部骨形成區的電鏡觀察顯示,除了成骨細胞在此發揮成骨作用外,成纖維細胞確實也存在着類似的成骨表現。例如,在其線粒體內可清晰見到鈣鹽顆粒,部分內質網腔內可見成熟的膠原纖維,分泌到其四周的膠原纖維內可見高密度的鈣鹽結晶沉積。不僅如此,成纖維細胞還能像成骨細胞一樣產生基質小泡並引起小泡內的鈣鹽沉積鈣化的基質小泡形成叢毛球狀的鈣球,鈣球隨後合併、融合為骨組織。以上種種現象表明,成纖維細胞與成骨細胞一樣具備提供鈣鹽沉積及骨形成所必需的條件。在從纖維性骨痂到骨性骨痂的演變過程中,成纖維細胞也隨之演變為骨細胞,與成骨細胞的歸宿相一致。但二者在演變過程中的表現又不盡相同,主要有以下幾點可資鑑別:
(1)成纖維細胞及其細胞核均呈不規則的橢圓形或長方形,而成骨細胞及其細胞核則為邊緣比較光整的橢圓形;
(2)成纖維細胞均單獨存在,細胞之間有眾多的膠原纖維相隔,成骨細胞則以連續排列的形式出現;
(3)成纖維細胞的細胞質內溶酶體少見,而成骨細胞的細胞質內則常有溶酶體可見;
(4)成纖維細胞四周的骨組織都由叢毛球狀鈣球或針狀鈣鹽結晶組成,成骨細胞則都有一面緊貼比較成熟與緻密的骨組織;
(5)成骨細胞是一個一個地被骨組織(類骨質)包圍變為骨細胞,而成纖維細胞則可以是兩個或兩個以上同時被骨組織包圍在一個陷窩內,然後再隨着細胞之間基質的鈣化而分隔為各占一個骨陷窩。
對成纖維細胞的成骨作用,有學者認為這是成纖維細胞的固有特性在骨折這一特定情況下得以表達的結果。骨折局部活的和失活的骨組織及軟骨組織,以及骨基質中的某些大分子都有可能誘導成纖維細胞表達其成骨作用進而演變為骨細胞。較早在骨基質中發現的骨形態發生蛋白( bone morphogenetic protein,BMP)即對成纖維細胞有一定的誘導作用對骨折癒合中BMP作用的研究,表明創傷使內源性BMP呈階段性合成與釋放,並誘導周圍軟組織中的間充質細胞或成纖維細胞等向成骨方向轉化。應用PAP法發現,骨折後第35天局部纖維肉芽組織中的成纖維細胞樣間充質細胞內以及第14天新生骨小梁間纖維組織中的成纖維細胞樣間充質細胞內,都與成骨細胞、軟骨細胞和骨基質一樣存在BMP,表明這些成纖維細胞樣間充質細胞已被誘導為可合成分泌BMP、具有成骨作用的細胞。而 Sampath從牛骨基質中分離提純得到的成骨素對成纖維細胞的骨誘導能力更是超過了BMP和當時已知的其他骨生長因子。
成纖維細胞在其成骨作用得以表達後,可能通過兩種方式成骨:(1)膜內成骨;(2)在環繞軟骨的纖維層內成骨。開始分泌膠原纖維後,參與成骨的成纖維細胞只有兩個歸宿:(1)變性、死亡、碎裂直至消失,這種演變發生早、範圍廣,故從纖維性骨痂形成開始,就逐漸有基質成分發生鈣化,進而轉變為骨基質;(2)演變為骨細胞,這一過程出現較晚,並穿插在前一過程之中,故在形成骨組織的細胞成分的同時,還使豐富的纖維骨痂演變為骨性骨痂形成骨組織。但這種由成纖維細胞演變成的骨細胞,其結局如何其生物學特性與由成骨細胞演化而來的骨細胞是否相同仍不清楚。例如,骨細胞從骨陷窩脫離後,可恢復為功能活躍的成骨細胞,再次參與骨組織的形成而由成纖維細胞演變成的骨細胞在脫離骨陷窩後,是成為成骨細胞還是恢復為成纖維細胞、此時是否還具備成骨作用等一系列問題尚缺乏研究。
參考文獻
- ↑ 纖維細胞的基本信息行業諮詢
- ↑ 什麼是成纖維細胞?_充質_合成_結締組織搜狐網