细胞骨架
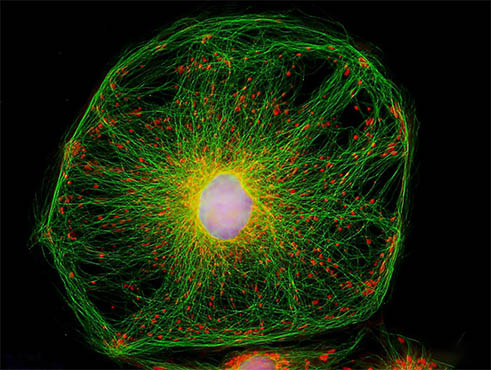 |
细胞骨架是真核细胞中的蛋白纤维网架体系。
狭义的细胞骨架(cytoskeleton)概念是指真核细胞中的蛋白纤维网架体系( 微管(microtubule, MT)、微丝(microfilament, MF)及中间纤维(intermediate filament, IF )组成的体系),[1]它所组成的结构体系称为“细胞骨架系统”,与细胞内的遗传系统、生物膜系统、并称“细胞内的三大系统”。直到20世纪60年代后,采用戊二醛常温固定,才逐渐认识到细胞骨架的客观存在,它是真核细胞借以维持其基本形态的重要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。广义的细胞骨架概念是细胞核骨架、细胞质骨架、细胞膜骨架和胞外基质所形成的网络体系。核骨架、核纤层与中间纤维在结构上相互连接,贯穿于细胞核和细胞质的网架体系。[2]
目录
作用
细胞骨架(cytoskeleton)是指真核细胞中的蛋白纤维网络结构,发现较晚,主要是因为一般电镜制样采用低温(0-4℃)固定,而细胞骨架会在低温下解聚。
直到20世纪60年代后,采用戊二醛常温固定,才逐渐认识到细胞骨架的客观存在。真核细胞借以维持其基本形态的重要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。
细胞骨架不仅在维持细胞形态,承受外力、保持细胞内部结构的有序性方面起重要作用,而且还参与许多重要的生命活动,如:在细胞分裂中细胞骨架牵引染色体分离,在细胞物质运输中,各类小泡和细胞器可沿着细胞骨架定向转运;在肌肉细胞中,细胞骨架和它的结合蛋白组成动力系统;在白细胞(白血球)的迁移、精子的游动、神经细胞轴突和树突的伸展等方面都与细胞骨架有关。另外,在植物细胞中细胞骨架指导细胞壁的合成。
微管
微管(microtubule)可在所有哺乳类动物细胞中存在,直径大于12nm,除了红细胞(红血球)外,所有微管均由约55kD的α及β微管蛋白(tubulin)组成。它们正常时以(αβ)二聚体形式存在,并以头尾相连的方式聚合,形成微管蛋白原纤维(protofilament),一般由13根这样的原纤维构成一个中空的微管,直径22~25nm。少数变异的微管如线虫等所有的则有其他数目的原纤维。微管确定膜性细胞器(membrane-enclosed organelle)的位置和作为膜泡运输的导轨。微管是细胞骨架的架构主干,并也是某些胞器的主体,例如中心粒(centriole)就是由9组3联微管组成的构造,而真核生物的纤毛(cilium)与鞭毛(flagellum)也是由以微管为9+2结构,即由9个二联微管和一对中央微管构成,其中二联微管由AB两个管组成,A管由13条原纤维组成,B管由10条原纤维组成,两者共享5条。A管对着相邻的B管伸出两条动力蛋白臂,并向鞭毛中央发出一条辐。基体的微管组成为9+0,并且二联微管为三联微管所取代。组成的轴丝(axoneme)为主体。
从各种组织中提纯微管蛋白可以发现还存在一些其他蛋白成分(5%-20%),称之谓微管相关蛋白(microtube associated proteins MAPs)。这些蛋白具有组织特异性,表现出从相同αβ二聚体聚合形成的微管具有独特的性质,已从人类不同组织中发现了多种α及β微管蛋白,并追踪微管基因表现出部分基因家族,某些基因被认为是编码独特的微管蛋白。
微管形成的有些结构是比较稳定的,是由于微管结合蛋白的作用和酶修饰的原因。如神经细胞轴突、纤毛和鞭毛中的微管纤维。大多数微管纤维处于动态的聚合和灾变(一种突然的,迅速的,一般不可逆转的分解)状态,这是实现其功能所必需的性质(如纺锤体)。与秋水仙素(colchicine)结合的微管蛋白可加合到微管上,并阻止其他微管蛋白单体继续添加,进而破坏纺锤体的结构,长春花碱具有类似的功能。紫杉酚(taxol),能促进微管的聚合,并使已形成的微管稳定,然而这种稳定性会破坏微管的正常功能。这些药物可以利用破坏微管功能以阻止细胞分裂,成为癌症治疗的新希望。
在人类至少发现两种明显区别的α-微管蛋白及三种明显区别的β-微管基因,它们产生具有特定功能的微管蛋白mRNA,由于这些编码在结构组分上十分近似蛋白质分子,在不同组织存在多少特异性的具有差异表达的微管蛋白亚型,尚待深入研究。
除了α-与β-微管蛋白有编码相似的不同变异型,近几年来又发现了多种编码差异更大的新的微管蛋白,形成不同的基因家族。其中gamma微管蛋白位于细胞内的微管组织中心(microtubule organizing center, MTOC),是用以提供α及β微管蛋白进行聚合反应形成微管的起始核心。而delta与epsilon则被认为与中心体(centrosome)的结构与形成有关。其他尚有eta, zeta, theta等等多种变异,不过通常仅存在少数几种真核单细胞生物如原虫或纤毛虫里,可能跟这些生物独特的结构与生理习性有关,进一步详情仍待研究 。
微丝
微丝(microfilament)也普遍存在于所有真核细胞中,是一个实心状的纤维,直径为4nm-7nm一般细胞中含量约占细胞内总蛋白质的1%-2%,但在活动较强的细胞中可占20%-30%。在一般细胞主要分布于细胞的表面,直接影响细胞的形状。微丝具有多种功能,在不同细胞的表现不同,在肌细胞组成粗肌丝、细肌丝,可以收缩(收缩蛋白),在非肌细胞中主要起支撑作用、非肌性运动和信息传导作用。
微丝主要由肌动蛋白(actin)构成,和肌球蛋白(myosin,一种分子马达蛋白)一起作用,使细胞运动。它们参与细胞的变形虫运动、植物细胞的细胞质流动与肌肉细胞的收缩:
植物细胞的细胞质流动: 微丝中的actin(肌动蛋白)与myosin(肌球蛋白)在细胞质形成三维的网络体系。actin位于外质,myosin位于内质。 myosin连结着细胞质颗粒,由ATP供给能量,myosin与细胞质颗粒的结合体沿着actin filament滑动,从而带动整个细胞质的环流。
变形虫运动(amoeboid movememt,阿米巴运动): 肌肉细胞的收缩:
如同微管蛋白,肌动蛋白的基因组成一个超家族,并组成多种极为相似的结构。例如,各种肌肉细胞有不同的肌动蛋白:①骨骼肌的条纹纤维;②心肌的条纹纤维;③血管壁的平滑肌;④胃肠道壁的平滑肌。它们在氨基酸组分上有微小的差异(大约在400个氨基酸残基序列中有4-6个变异),在肌肉与非肌细胞中都还存在β及γ肌动蛋白,它们与具有横纹的α肌动蛋白可有25个氨基酸的差异。
G-肌动蛋白单体(含ATP)可聚合为呈纤维状的F-肌动蛋白(含ADP),它们可由Mg2+及高浓度的K+或Na+诱导而聚合,聚合后ATP水解为ADP及C-肌动蛋白ADP单体,组成F-肌动蛋白。在骨骼肌的细肌丝(thin filament,由肌动蛋白构成)与粗肌丝(thick filament,由肌球蛋白构成)相互作用而使肌肉收缩(肌球蛋白可以起作肌动蛋白激活的ATPase的作用)。肌球蛋白也存在于哺乳动物的非肌细胞中(但以非聚合状态存在)。
中间纤维
细胞骨架的第三种纤维结构称中间纤维(intermediate filament,IF),又称中间丝、中等纤维,直径介于微管和微丝之间(8nm-10nm),其化学组成比较复杂。构成它的蛋白质多达5种,常见的有波形蛋白(vimentin)、角蛋白(keratin)、结蛋白、神经元纤维、神经胶质纤维。在不同细胞中,成分变化较大。中间纤维使细胞具有张力和抗剪切力。中间纤维有共同的基本结构,即构建成一个中央α螺旋杆状区,两侧则是大小和化学组成不同的端区。端区的多样性决定了中间纤维外形和性质的差异和特异性。
以上这些结构单元并非是一成不变的,而是随细胞的生命活动而呈现高度的动态性,它们均由单体蛋白以较弱的非共价键结合在一起,构成纤维型多聚体,很容易进行组装和去组装,这正是实现其功能所必需的特点。
原核生物
长期以来,人们认为细胞骨架仅为真核生物所特有的结构,但近年来的研究发现它也存在于细菌等原核生物中。
人们已经在细菌中发现的FtsZ、MreB 和CreS 依次与真核细胞骨架蛋白中的微管蛋白、肌动蛋白丝及中间丝类似。FtsZ 能在细胞分裂位点装配形成Z 环结构,并通过该结构参与细胞分裂的调控;MreB能形成螺旋丝状结构,其主要功能有维持细胞形态、调控染色体分离等;CreS存在于新月柄杆菌中,它在细胞凹面的细胞膜下面形成弯曲丝状或螺旋丝状结构,该结构对维持新月柄杆菌细胞的形态具有重要作用。
疾病及危害
细胞骨架是细胞生命活动中不可缺少的细胞结构,其形成的复杂网络体系对细胞形态的改变和维持、细胞的分裂与分化、细胞内物质运输、细胞信息传递、基因表达等均具有重要意义。肿瘤、许多遗传性疾病、某些神经系统疾病等的发生均与细胞骨架的异常有关。临床上,常利用细胞骨架在不同细胞内的特异性分布的特征,来诊断某些疑难疾病,并依据细胞骨架与疾病的关系来设计并指导用药,开展防病治病。
细胞在病理情况下常常会出现细胞骨架系统异常。如阿尔茨海默症患者,在脑神经元中发现有大量扭曲变形的微管和大量受损的中间纤维;在恶性转化的细胞中,常表现为微管减少和解聚,细胞骨架异常可增强癌细胞的运动能力。研究表明,微丝束及其末端黏着斑的破坏以及肌动蛋白小体的出现,与肿瘤细胞的浸润和转移特性有关。
此外,中间纤维的分布具有严格的组织特异性,绝大多数肿瘤细胞在发生转移后仍表现其原发肿瘤的中间纤维类型,故可作为临床肿瘤的鉴别诊断和肿瘤细胞是否转移的判据。中间纤维显微技术与羊膜刺穿结合,可用于先天胎儿畸形的诊断,例如,若羊水中含有神经元纤维和神经胶质纤维细胞,则提示胎儿或有中枢神经系统畸形。
肿瘤
(一)细胞骨架在肿瘤细胞中的变化
机体中各组织细胞的结构和功能是密切相关的,细胞骨架无论在组装还是分布上若发生了变化,必将影响到细胞的功能。在恶性转化的细胞中,常表现为细胞骨架结构的破坏、组装和分布的异常、微管的解聚等。
我国学者对胃癌、鼻咽癌、食管癌、肺鳞癌、肺小细胞癌、肺腺癌、小鼠肉瘤等9株肿瘤细胞进行观察,发现肿瘤细胞质内免疫荧光染色的微管减少甚至缺如,且癌细胞的微管变化主要发生在间期,而在分裂期,纺锤体微管与正常细胞相同。用荧光抗体技术已证明,在长期传代的癌变细胞内微管显著减少。因此,微管数量的减少是细胞恶性转化的重要标志。而且肿瘤细胞内原有的微丝束明显减少甚至消失,常出现肌动蛋白凝聚小体;肿瘤细胞内的微丝分布异常,无序紊乱,常不与细胞膜相连。
肿瘤细胞的浸润转移是一个极其复杂的过程,在此过程中,某些细胞骨架成分的改变可增强癌细胞的运动能力。研究表明,微丝束和其末端黏着斑的破坏以及肌动蛋白小体的出现,可能与肿瘤浸润转移的特性有关,肌动蛋白小体形成,可能代表肿瘤细胞高转移的恶性表型。
(二)中间纤维与肿瘤诊断
中间纤维形态相似,但具有严格的组织特异性,不同类型的中间纤维严格地分布在不同类型的细胞中,故可根据中间纤维的种类区分上皮细胞、肌肉细胞、间质细胞、胶质细胞和神经细胞。在这几种纤维类型的基础上,中间纤维还可进一步分出若干亚型。因绝大多数肿瘤细胞在生长时,继续保持其来源细胞的中间纤维的种类、超微结构和免疫学特性,如癌是以上皮细胞的角质蛋白为特征的,肌肉瘤是以结蛋白、非肌肉瘤是以波形纤维蛋白、神经胶质瘤是以神经胶质纤维酸性蛋白、从交感神经来的肿瘤是以神经纤维蛋白(necotinamide protein)为特征的。所以,人们可根据中间纤维的种类,来鉴别、区分不同组织来源的肿瘤细胞及各肿瘤细胞的亚型,为肿瘤的诊断和治疗提供决定性的依据。
在临床上,中间纤维蛋白的荧光素标记抗体技术在细胞分类尤其是肿瘤细胞的鉴别上,具有广泛的应用前景。如采用角蛋白荧光素标记抗体确认上皮癌,用波形蛋白荧光素标记抗体确认淋巴肉瘤、黑色素瘤、骨肉瘤等,用结蛋白荧光素标记抗体确认肌细胞肉瘤,用神经纤维蛋白荧光素标记抗体确认神经母细胞瘤、神经节母细胞瘤、嗜铬细胞瘤等。
(三)微管和微丝与抗肿瘤药物
在有丝分裂的中/后期,秋水仙素和长春花碱等化合物可与纺锤体微管蛋白或微管结合,抑制细胞增殖。研究表明具有抗有丝分裂能力的药物如紫杉醇和长春新碱等抑制细胞增殖和杀死肿瘤细胞的主要机制是稳定纺锤体微管动力学,而不是使微管解聚或过度多聚化,在有丝分裂的中/后期抑制细胞分裂,诱导细胞凋亡。
细胞松弛素是真菌的代谢产物,也是研究最多且应用最广泛的微丝特异性工具药,它作用于肌动蛋白,可与微丝正端结合,抑制其聚合,使微丝解聚,导致细胞表面皮质层松解,引起细胞表面起泡,使微绒毛变成茬状物,细胞整体形态呈现树枝状化,并可抑制各种依赖于微丝的运动,具有抗肿瘤潜能。
神经系统疾病
阿茨海默病(Alzheimer’s disease,AD)即早老性痴呆病,属微管遗传性疾病。对患者脑脊液分析发现,AD患者脑脊液中tau蛋白含量明显高于非AD患者和正常人,且其神经元中还可见到大量损伤的神经元纤维,它们由成对的螺旋状纤维(paired helical filament,PHF)和相对较直的纤维(straight filament,SF)组成,主要成分是高磷酸化状态的tau蛋白,其性能稳定。对死亡AD患者的大脑进行分析发现,神经元中微管蛋白的数量并无异常,但微管聚集缺陷。孤立的微管蛋白与结合的微管蛋白均可以高磷酸化的方式与其他配体结合形成稳定的tau蛋白。因为微管是轴浆流必需的细胞骨架,因此AD中微管聚集缺陷,可使微管聚合发生障碍,微管扭曲变形,可能引起轴浆流阻塞,导致神经元纤维包涵体的形成,从而使神经信号传递紊乱,影响轴质的物质运输,使神经元的营养和代谢产生障碍,从而出现痴呆现象。
Tau蛋白及其他一些细胞骨架蛋白的异常还可引起其他神经系统疾病,如运动神经元疾病、帕金森病、肌强直性营养不良等。
遗传性疾病
某些遗传性疾病常与细胞骨架的异常或细胞骨架蛋白基因的突变有关。WAS (Wiskoff-Aldrich syndrome)是一种遗传性免疫缺陷疾病,其特征是湿疹、出血和反复感染。研究表明,微丝的异常是引起WAS的根源所在。
随着研究方法和手段的不断改进,尤其是利用转基因小鼠或基因敲除小鼠进行研究,发现中间纤维与许多遗传疾病有关。人类遗传性皮肤病单纯性大疱性表皮松解症(epidermolysis bullosasimplex,EBS)是最典型的例证,该病是由角蛋白14(CK14)基因突变所致。
衰老
老年病学研究表明,老年人随着年龄的增加,机体各细胞均出现功能低下的表现。这与细胞骨架的数量、结构及功能的变化有关。动物实验表明,老龄动物的神经元内微管数量减少,腹腔巨噬细胞内的微丝数量减少,可影响神经信号传递,影响轴质的物质运输,影响神经元的营养和代谢,影响免疫机能,进而影响到细胞的功能。所以,老年人的衰老表现为脑功能衰退和机体免疫等多系统多功能的低下。
其他蛋白
不仅如此,细胞骨架还包含有很多结构单元的附属蛋白质,比如:
分子马达(molecular motors): 动力蛋白(dynein), kinesin, myosin 结合蛋白:vinculin, cofilin, tropomyosin等等
广义的细胞骨架还包括核基质(nucleoskeleton)、核纤层(nuclear lamina)和细胞外基质(extracellular matrix),形成贯穿于细胞核、细胞质、细胞外的一体化网络结构。
发现历史
细胞骨架(cytoskeleton)是指真核细胞中的蛋白纤维网络结构。发现较晚,主要是因为一般电镜制样采用低温(0-4℃)固定,而细胞骨架会在低温下解聚。
直到20世纪60年代后,电镜制样采用戊二醛进行常温固定,人们才逐渐认识到细胞骨架的客观存在。真核细胞借以维持其基本形态的重要结构,被形象地称为细胞骨架,它通常也被认为是广义上细胞器的一种。细胞骨架不仅在维持细胞形态,承受外力、保持细胞内部结构的有序性方面起重要作用,而且还参与许多重要的生命活动,如:在细胞分裂中细胞骨架牵引染色体分离,在细胞物质运输中,各类小泡和细胞器可沿着细胞骨架定向转运;在肌肉细胞中,细胞骨架和它的结合蛋白组成动力系统;在白细胞(白血球)的迁移、精子的游动、神经细胞轴突和树突的伸展等方面都与细胞骨架有关。另外,在植物细胞中细胞骨架指导细胞壁的合成。
参考文献
- ↑ 细胞生物学 | 细胞骨架_蛋白质_纤维_中心搜狐网
- ↑ 什么是细胞骨架?-资讯分析测试百科网wiki版