纤维细胞查看源代码讨论查看历史
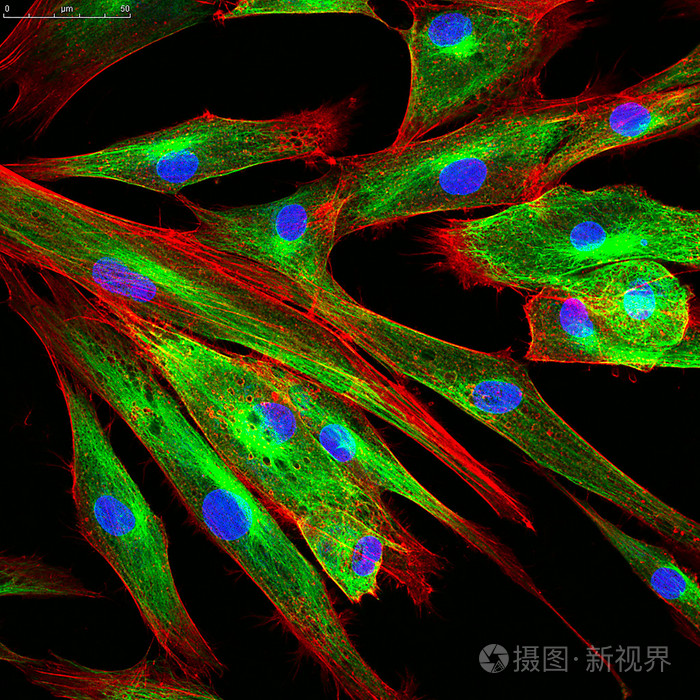 |
纤维细胞(fibrocyte)是机能不活跃的成纤维细胞。胞体呈梭形,胞质较少,弱嗜酸性,胞核小,染色深。电镜下可见,细胞中粗面内质网和高尔基体均不发达。当组织受损时,纤维细胞可转化为成纤维细胞而参与修复过程。[1]
简介
纤维细胞和成纤维细胞是处于不同功能状态的同一种细胞,如在组织损伤后的修复过程中,纤维细胞可转化为功能活跃的成纤维细胞。纤维细胞较小,呈多角形,胞质较少,弱嗜酸性,胞核亦较小,染色较深,电镜下粗面内质网较少,高尔基复合体不发达。成纤维细胞较大,呈星形或梭形,有突起,细胞核呈卵圆形,染色质稀疏,染色淡,核仁明显,胞质较多。成纤维细胞能合成和分泌胶原蛋白、弹性蛋白和蛋白多糖,形成胶原纤维、弹性纤维和网状纤维以及基质成分。[2]
转变
在结缔组织中,成纤维细胞还以其成熟状态—纤维细胞(fibrocyte)的形式存在,二者在一定条件下可以互相转变。 不同类型的结缔组织含成纤维细胞的数量不同。通常,疏松结缔组织中成纤维细胞的数量比同样体积的致密结缔组织中所含成纤维细胞的数量要少,故分离培养成纤维细胞多以真皮等致密结缔组织为取材部位。
形态
成纤维细胞形态多样,常见的有梭形、大多角形和扁平星形等,其形态尚可依细胞的功能变化及其附着处的物理性状不同而发生改变。成纤维细胞胞体较大,胞质弱嗜碱性,胞核较大呈椭圆形,染色质疏松着色浅,核仁明显。电镜下,其胞质可见丰富的粗面内质网、游离核糖体和发达的高尔基复合体,表明它具有合成和分泌蛋白质的功能。成纤维细胞尚可合成和分泌胶原纤维、弹性纤维、网状纤维及有机基质。它合成的前胶原蛋白分子经内切酶作用,聚合和重排,可形成与成骨细胞合成分泌的胶原原纤维一样具有64nm(640?)周期横纹的胶原原纤维,胶原原纤维经互相粘合形成胶原纤维。经检测,这两种细胞合成分泌的胶原纤维均是Ⅰ型胶原纤维,在形态和生化结构上完全相同。 处于成熟期或称静止状态的成纤维细胞,胞体变小,呈长梭形,粗面内质网和高尔基复合体均不发达,被称为纤维细胞。在外伤等因素刺激下,部分纤维细胞可重新转变为幼稚的成纤维细胞,其功能活动也得以恢复,参与组织损伤后的修复。另外,在结缔组织中,仍保留着少量具有分化潜能的间充质细胞,它们在创伤修复等情况下可增殖分化为成纤维细胞。
作用
创伤修复
创伤均会造成不同程度的细胞变性、坏死和组织缺损,必须通过细胞增生和细胞间基质的形成来进行组织修复。在此修复过程中,成纤维细胞起着十分重要的作用。以伤口愈合过程为例,成纤维细胞通过有丝分裂大量增殖,并从4~5天或6天开始合成和分泌大量的胶原纤维和基质成分,与新生毛细血管等共同形成肉芽组织,填补伤口组织缺损,为表皮细胞的覆盖创造条件。在伤口愈合中,成纤维细胞主要来源于真皮乳头层的局部成纤维细胞和未分化的间充质细胞,以及血管周围的成纤维细胞和周细胞内脏损伤时,参与修复过程的成纤维细胞多来自间质和包膜,以及粘膜下或浆膜下层的结缔组织。有人认为创伤愈合过程中伤处聚集的大量成纤维细胞,一方面是由成纤维细胞通过分裂增殖而来,另一方面,更多地是由邻近的间充质细胞、纤维细胞和毛细血管周细胞等演变或游走到伤处。在创伤修复的后期,成纤维细胞通过分泌胶原酶参与修复后组织的改建。在某些病理条件下,以成纤维细胞为主要细胞成分的肉芽组织或增生组织块还可以在非骨组织内发生钙化,引起异位骨化( ectopicossification)。但对于异位骨化的参与细胞及其机制尚不十分清楚,未分化间充质细胞、成纤维细胞、内皮细胞和毛细血管周细胞等可划归为诱导性骨祖细胞的细胞都有可能参与这一过程。
骨创伤修复
最简单和常见的骨创伤即是骨折,其愈合过程须经过炎性反应、清扫、纤维骨痂和骨性骨痂4个阶段。不同阶段参与的细胞主体不同。成纤维细胞从骨折第3天起就出现于骨折局部血肿中,骨折后5天即在机化血肿及骨折断端的间隙及其周围大量存在,是参与纤维骨痂阶段的主要细胞成分。在此阶段成纤维细胞一方面大量分裂增殖,一方面又合成和分泌大量I型胶原,使肉芽组织逐步变成疏松的结缔组织,将骨断端包围起来,形成接合两骨折断端的巨大的纤维骨痂然而,这种由无数成纤维细胞和丰富的肉芽组织为主体构成的纤维结缔组织却不会演变为在其他组织创伤修复时常见的癜痕组织,而是通过钙盐结晶在其内部不断沉积,逐渐演变为骨性骨痂,使骨折局部的修复达到骨性愈合,恢复骨组织的结构。此时,骨折愈合部只有骨组织而不再存在成纤维细胞。
成骨作用
对骨折局部骨形成区的电镜观察显示,除了成骨细胞在此发挥成骨作用外,成纤维细胞确实也存在着类似的成骨表现。例如,在其线粒体内可清晰见到钙盐颗粒,部分内质网腔内可见成熟的胶原纤维,分泌到其四周的胶原纤维内可见高密度的钙盐结晶沉积。不仅如此,成纤维细胞还能像成骨细胞一样产生基质小泡并引起小泡内的钙盐沉积钙化的基质小泡形成丛毛球状的钙球,钙球随后合并、融合为骨组织。以上种种现象表明,成纤维细胞与成骨细胞一样具备提供钙盐沉积及骨形成所必需的条件。在从纤维性骨痂到骨性骨痂的演变过程中,成纤维细胞也随之演变为骨细胞,与成骨细胞的归宿相一致。但二者在演变过程中的表现又不尽相同,主要有以下几点可资鉴别:
(1)成纤维细胞及其细胞核均呈不规则的椭圆形或长方形,而成骨细胞及其细胞核则为边缘比较光整的椭圆形;
(2)成纤维细胞均单独存在,细胞之间有众多的胶原纤维相隔,成骨细胞则以连续排列的形式出现;
(3)成纤维细胞的细胞质内溶酶体少见,而成骨细胞的细胞质内则常有溶酶体可见;
(4)成纤维细胞四周的骨组织都由丛毛球状钙球或针状钙盐结晶组成,成骨细胞则都有一面紧贴比较成熟与致密的骨组织;
(5)成骨细胞是一个一个地被骨组织(类骨质)包围变为骨细胞,而成纤维细胞则可以是两个或两个以上同时被骨组织包围在一个陷窝内,然后再随着细胞之间基质的钙化而分隔为各占一个骨陷窝。
对成纤维细胞的成骨作用,有学者认为这是成纤维细胞的固有特性在骨折这一特定情况下得以表达的结果。骨折局部活的和失活的骨组织及软骨组织,以及骨基质中的某些大分子都有可能诱导成纤维细胞表达其成骨作用进而演变为骨细胞。较早在骨基质中发现的骨形态发生蛋白( bone morphogenetic protein,BMP)即对成纤维细胞有一定的诱导作用对骨折愈合中BMP作用的研究,表明创伤使内源性BMP呈阶段性合成与释放,并诱导周围软组织中的间充质细胞或成纤维细胞等向成骨方向转化。应用PAP法发现,骨折后第35天局部纤维肉芽组织中的成纤维细胞样间充质细胞内以及第14天新生骨小梁间纤维组织中的成纤维细胞样间充质细胞内,都与成骨细胞、软骨细胞和骨基质一样存在BMP,表明这些成纤维细胞样间充质细胞已被诱导为可合成分泌BMP、具有成骨作用的细胞。而 Sampath从牛骨基质中分离提纯得到的成骨素对成纤维细胞的骨诱导能力更是超过了BMP和当时已知的其他骨生长因子。
成纤维细胞在其成骨作用得以表达后,可能通过两种方式成骨:(1)膜内成骨;(2)在环绕软骨的纤维层内成骨。开始分泌胶原纤维后,参与成骨的成纤维细胞只有两个归宿:(1)变性、死亡、碎裂直至消失,这种演变发生早、范围广,故从纤维性骨痂形成开始,就逐渐有基质成分发生钙化,进而转变为骨基质;(2)演变为骨细胞,这一过程出现较晚,并穿插在前一过程之中,故在形成骨组织的细胞成分的同时,还使丰富的纤维骨痂演变为骨性骨痂形成骨组织。但这种由成纤维细胞演变成的骨细胞,其结局如何其生物学特性与由成骨细胞演化而来的骨细胞是否相同仍不清楚。例如,骨细胞从骨陷窝脱离后,可恢复为功能活跃的成骨细胞,再次参与骨组织的形成而由成纤维细胞演变成的骨细胞在脱离骨陷窝后,是成为成骨细胞还是恢复为成纤维细胞、此时是否还具备成骨作用等一系列问题尚缺乏研究。
参考文献
- ↑ 纤维细胞的基本信息行业咨询
- ↑ 什么是成纤维细胞?_充质_合成_结缔组织搜狐网