木質部檢視原始碼討論檢視歷史
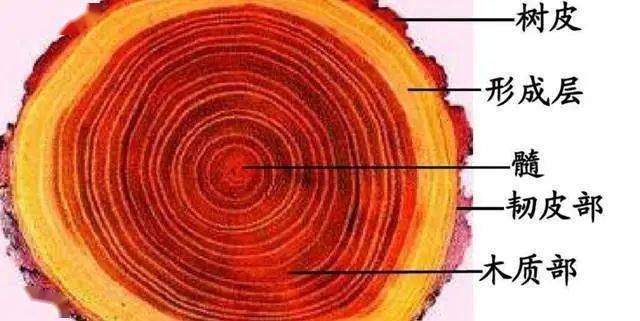 |
木質部是樹幹的主要組成部分,具有重要的經濟價值,[1]在所謂的「硬木」或雙子葉木本樹種中,木質部由導管、管胞、木纖維和木薄壁組織細胞以及木射線組成,常與韌皮部結合組成維管束,分布在植物體內形成維管系統[2]
定義
木質部指維管植物體內具有輸導和機械作用的一種複合組織。由導管、管胞、木纖維和木薄壁細胞等組成,常與韌皮部結合組成維管束,分布在植物體內形成維管系統。導管和管胞為輸導水和溶於水的無機鹽的管狀分子,為木質部的主要組成部分。
分類介紹
導管
導管( vessel)是由多個長管狀的死細胞縱向連接而形成的管道,其中每個細胞稱為導管分子( vessel member,vessel element)。在導管的形成過程中,每一細胞的橫壁逐漸全部溶解而形成單個的大孔隙,或若干個局部溶解而形成數個較小的孔隙。在孔隙處無任何細胞壁的壁層殘留,這種孔隙稱為穿孔(perforation)。具有穿孔的橫壁稱為穿孔板( perforation plate)。其中,只具1個穿孔的稱單穿孔板,具2個以上穿孔的稱復穿孔板。穿孔的形成使導管中導管分子問的鄰接壁(即橫壁)打通,整個導管成為一個貫通的管道,因此其輸水的速度遠比管胞為快。此外,導管還可通過其側壁上無次生壁的部位與相鄰的其他細胞進行水分的橫向輸導。導管的次生壁也像管胞的次生壁那樣形成紋理。紋理形成的原因,是次生壁在有些地方鋪蓋,在另一些地方不鋪蓋,同時這兩種地方成互相間隔或穿插式地分布。根據所形成的紋理之不同,將導管分為下列類型。
1.環紋導管(annular vessel)
其次生壁沿着管狀初生壁的內表面,形成多個間隔一定距離的環。其管徑較小,存在於植物幼嫩器官中,如玉蜀黍、半夏等植物的幼莖中。
2. 螺紋導管(spiral vessel)
其次生壁從管狀初生壁的內表面的一端,成螺旋的帶狀鋪向另一端。其管徑與環紋導管的管徑差不多或者大一些,也多存在於植物的幼嫩器官中,如蓮藕(根莖)和豆芽的莖中。
3. 梯紋導管(scalariform vessel)
其次生壁既橫向地形成許多個間隔較短距離的環,又縱向地形成若干條長而直的帶,並與前者交連。管壁上無次生壁的地方成為許多個呈扁圓形或長方形的單紋孔,這些單紋孔沿縱軸方向排成若干條整齊的、互不交錯的隊列。這種導管的直徑稍大於上述兩種導管的直徑,多存在於植物的成長器官中,如葡萄的莖、荊三棱的球莖、香附的塊莖中。
4. 網紋導管(reticulate vessel)
其次生壁既作一些橫向的條狀鋪蓋,也作一些斜向的條狀鋪蓋,有時還作一些縱向的條狀鋪蓋;這些條狀次生壁短的多長的少,相互交連;無次生壁的地方成為一些等大或不等大、同形(但不會都呈圓形)或不同形的坑(單紋孔),它們之間多少呈交錯排列,且離得較近。這種導管的管徑較大,多存在於器官的成熟部分,如大黃的根及根莖、南瓜的莖和前胡的根中。
5. 孔紋導管(pitted vessel)
其導管壁上無次生壁的地方為一些具緣紋孔或直徑較小且呈圓形的單紋孔。其管徑較大,多存在於器官的成熟部分,如甘草的根、威靈仙的根和根莖中。
以上所述為5種紋理很典型的導管。在實際的觀察中常可以在同一導管上見到呈兩種紋理的次生壁,如環紋和螺紋次生壁,梯紋和網紋次生壁等。
某些植物的導管在發育過程中,其鄰接的薄壁細胞發生膨脹,進而通過導管壁上無次生壁的部位或紋孔侵入導管腔內,形成大小不等的囊泡狀突起。
管胞
管胞( tracheid)是絕大多數蕨類植物和裸子植物的輸導組織,同時兼有支持作用。有些被子植物或被子植物的某些器官中也有管胞,但不是主要的輸導組織。
管胞中的每個細胞,全形呈兩端尖的長管狀,成熟後無原生質體,細胞壁含次生壁層且木質化。次生壁形成各種紋理,因而有環紋管胞、螺紋管胞、梯紋管胞、孔紋管胞等類型。管胞的管徑較導管的小,且水分從一個管胞細胞運行到另一個管胞細胞的方式,是從紋孔對處滲透過去,因此其運輸水分的效率較導管低,是一類原始的輸水組織。管胞的主要類型有5種:環紋管胞、螺紋管胞、梯紋管胞、孔紋管胞、紋孔。
木纖維
雙子葉木本植物的次生木質部內的纖維。細胞壁厚,細胞腔較窄,形體細長,木質化,是一種厚壁組織,有退化程度不同的具緣紋孔。木纖維有兩種主要類型:(1)纖維管胞,具有具緣紋孔;(2)韌型纖維,細胞壁較厚,具單紋孔。木纖維主要起支持作用,有貯藏和輸導作用。
木薄壁組織細胞
在次生木質部中一般可分為軸向薄壁組織細胞和徑向薄壁組織細胞(即「木射線」)兩種。其細胞成熟後,一般都具有加厚的次生壁,並木質化。如有次生壁,則厚壁組織細胞壁之間的紋孔對有具緣、半具緣的類型。木薄壁組織細胞主要起貯藏作用。在木材橫切面上,依據軸向薄壁組織細胞與導管相對位置的不同,可分為離管薄壁組織和傍管薄壁組織兩大類。
木射線
位於形成層以內次生木質部中的維管射線。其主要功能是橫向運輸和貯藏。在橫切面上,木射線呈輻射狀排列。在切向切面上,射線呈縱線或紡錘形。在徑向切面上,射線呈不同高度的線狀或片狀排列。木射線單列或多列、高度從一層到多層細胞。小的射線也可聚合成群,形成聚合射線。組成木射線的薄壁組織細胞,依據細胞最長軸方向的不同,可分為徑向伸長的橫臥細胞,以及方形或沿縱向伸長的直立細胞兩種類型。如果由同一種類型細胞組成的射線,或僅由射線薄壁組織細胞組成的,稱為同型射線,由不同類型的薄壁組織細胞組成的射線(雙子葉植物)或在裸子植物中由木薄壁組織細胞和射線管胞構成的射線,稱異型射線。
因此我們把樹幹或枝幹的木質看做是由龐大的連鎖的微觀細胞叢組成的,其中有些在器官功能成熟後變得高度木質化並死去;其他與此交織在一起的成分(組成術質部的薄壁組織和放射線)仍然存活,而且代謝活動的程度不同,所有這些細胞來源於精細的分生(周期性分裂)細胞柱體,術語叫做維管形成層,它們形成術質部和樹皮之問的分界線毋庸置疑,維管形成層作為一種分生組織或者形成組織,不僅在樹的發育過程中發揮重要的作用,而且在樹的整個生命歷程中都具有重要的作用。
特徵
木質部細胞的壁多數木質化,故有一定的機械支持作用。由莖與根的近端部的原形成層分化形成初生木質部,多年生木本植物由於維管形成層的逐年活動,不斷增長次生木質部,從而長成粗壯的樹幹。
木質部解剖學家習慣研究3種薄切面:橫切面、徑向切面和切向切面。徑向切面是通過莖的中心(直徑)所作的縱切面,切向切面是垂直於莖的半徑所作的縱切面。從微觀上看這3種方式結合在一起足以提供鑑別木質部所需要的所有信息:在大多數情況下,可以鑑別出木質所從屬的屬,有時甚至能鑑別其所屬種類。雖然通過該過程我們可以了解木質部許多特點,但是還有其他可利用的特徵,不需要顯微鏡就可以檢測到,其中包括一些結構特徵用放大鏡或者憑肉眼就可以看到,例如顏色和比重,以及樹皮的形態。在分析木質部顏色的時候,通常必須考慮活的外層區域和通常沒有活性的核心區域的差異——前者一般是顏色較淺的邊材,而後者是顏色較深的心材。從商業角度來看,心材是最有價值的,它裡面包含有數量不等的樹膠、樹脂和作為天然防腐劑的沉積物。對於有些木材,這種心材具有與眾不同的顏色,例如黑檀樹的是黑色的,豆科的洋蘇木樹的是紫色的。
木材的比重差異很大,正是這一重要的物理特徵使得人們可以據此分類:柚木是價值不菲的木材,木質密實、厚重,並且由於裡面有樹脂材料沉積所以特別不易腐爛;與此相反,美國香脂木特別輕,有點類似於分布比較廣泛的西非樹,有一定的價值。毋庸置疑,樹的用途與顯微鏡下可觀察到的它的解剖結構密不可分。
木質素
在樹木生長以及存活的時候,樹的導管壁、管胞壁和纖維壁充滿一種叫做木質素的物質:當它們死亡後,這種物質仍然存在,因此保留了硬的細胞形狀。
木質素是一系列化學成分不詳的複合碳化合物的通用術語,但是很明顯有許多不同種類的木質素。一個值得一提的有趣事實是針葉樹的木質素與闊葉樹的木質素不同。另外我們需要注意的重要事實是:這些細胞和它們的木質素決定木質的強度和硬度,因此木質素壁越厚.這些特性就越明顯。
生長
維管植物的初生生長和分化時,由原形成層分化的木質部,稱為初生木質部,其中含有纖維和薄壁組織細胞,但沒有射線,所以,不構成軸向系統和徑向系統。在莖、葉和花等部位中,初生木質部和初生韌皮部結合成束狀,即為維管束。在莖的維管束之間有薄壁組織,稱為髓射線。在根中,初生木質部在中央形成一個核心,核心中央具髓或不具髓。根據分化的早晚,初生木質部又可分為原生木質部和後生木質部兩部分。
原生木質部
木質部中最早形成的部分。在莖、葉中,原生木質部是在活躍伸長的組織中成熟的,因此會受到引力的影響,其中無生命的管狀分子被拉伸而毀壞。在根中,原生木質部管狀分子成熟的地方,是在伸長區以後,所以存在時間較久。原生木質部一般只有管狀分子及其包圍它們的薄壁組織。原生木質部的管狀分子,主要有環紋和螺紋加厚。
後生木質部
初生木質部中,原生木質部分化以後所形成的部分,如有次生木質部時,則在次生木質部發生之前產生。後生木質部一般是在初生植物體仍舊生長時開始發生,其成熟則是在伸長完成以後,因此,比原生木質部受周圍組織伸長生長的影響小。後生木質部的組成比原生木質部複雜,除了管狀分子和薄壁組織外,還有纖維。薄壁組織可以散布在管狀分子之中,也可以形成射線狀的徑向行列。在切向切面上,它們是軸向的薄壁組織細胞。後生木質部的管狀分子可以有環紋、螺紋、梯紋以及具緣紋孔等。
次生木質部
維管植物次生生長時,由維管形成層產生的木質部。次生木質部分為軸向系統和徑向或射線系統。軸向系統指次生維管組織中,由紡錘狀形成層原始細胞衍生的細胞,它們的長軸與植物體軸呈平行排列。徑向系統是次生維管組織中所有射線的總稱。其細胞的長軸與體軸間成水平方向排列。
結構
年輪
人們都知道在攔腰劈開樹幹之後會看到年輪,這种放射狀的生長輪代表着它的生長年齡,但是生長年輪是怎樣出現的呢?答案就在於維管形成層在每年不同時期產生的細胞類型和大小。在有些例種中,年輪看起來更加明顯,這是因為春季初次形成的導管比夏季形成的導管要大得多,英國橡樹刪岑樹部很好地表現這種特徵,這類木質稱為「環孔材」,有些木材,包括山毛櫸樹、柳樹、蘋果樹等,在春季和夏季木質之間導管直徑沒有明顯的差異,這類術質的年輪叫做「散孔材」,其年輪問的界線並不十分清晰——夏季這類木質除了導管尺寸外,還存在其他明顯的特徵作為分界標記。在種軟木中,例如蘇格蘭松,在生長周期結束時形成管胞,此時正是夏季,它們比起春季形成的管胞具有更窄的孔和更厚的壁,由此人們能夠識別每年生長的輪。
單子葉的被子植物例如棕櫚一般沒有顯示出由於維管形成層柱所致的每年樹幹的變粗,也就是說沒有明顯的次生增厚過程。但是有極少數單子葉被子植物樹幹變粗的方式非常怪,完全不同於雙子葉植物,最著名的例子就是龍血樹,它的基本組織里的細胞是分生的―能夠像我們前面描述的雙子葉植物的形成層細胞那樣分裂,但是它不是向外莖部切掉韌皮部細胞,也不是向着中心切斷木質部細胞,而是韌皮部細胞和木質部細胞都向着莖的中心被切掉,這樣我們會看到如同在初生莖里維管束那樣的模式不斷複製、增加。
疊生木材和非疊生木材
在種子植物木材中,軸向細胞和射線在切向切面上排列成同一水平方向,稱為疊生木材。疊生木材是由疊生形成層所產生。例如,刺槐屬的木材。非疊生木材是由非疊生形成層產生的,木材中軸向細胞和射線細胞在切向切面上並不排列成水平系列。例如胡桃屬的木材。
邊材和心材
根和莖的木材外部,含有生活的細胞和貯存物質、並具輸導水分和無機鹽功能的部分,稱為邊材。在次生木質部內部已經停止貯藏和輸導作用的部分,稱心材,其顏色一般比邊材深。邊材的數量較為恆定,心材的相對數量在各種植物中都不同,而且還受環境的影響。一般說來,心材的數量可以不斷增加,在少數植物中,也可不形成心材。
軟材與硬材
軟材指裸子植物,特別是松柏類植物的木材。裸子植物木材的結構比較簡單,大多數種類最明顯的特徵是沒有導管,主要由管胞組成。在軟材中,可以有纖維管胞,但是沒有韌型纖維。硬材指雙子葉植物的木材。雙子葉植物的木材結構比裸子植物的變化大。原始無導管的雙子葉植物木材結構比較簡單,但在具導管的雙子葉植物木材中,還有管胞、一種或幾種纖維、軸向薄壁組織細胞和一種或幾種射線。另外,在許多雙子葉植物木材中,當導管變得不活躍以後,與導管相鄰的軸向薄壁組織細胞和射線薄壁組織細胞,可以通過紋孔形成突起,填充在導管腔內,形成侵填體。
散孔材與環孔材
在雙子葉植物次生木質部的橫切面上,生長輪中的導管管孔分布相當均勻,或從早材到晚材的管孔大小逐漸變化,這種木材稱散孔材,例如毛白楊。環孔材是指雙子葉植物的次生木質部中,早材的導管管孔明顯地比晚材大,因而在橫切面上形成清楚的同心[[環狀排列。例如水曲柳。
應力木是莖或枝條在抵抗迫使它們傾斜或彎曲的重力作用下形成的木材。松柏類植物和雙子葉植物的應力木在產生部位與結構等方面都有相當大的差別,因此又可分為壓縮木和伸張木兩種。壓縮木,即松柏類植物的應力木。在這類植物中,傾斜或彎曲莖或枝條下側的木材與正常的木材相比,組織質地緻密,木質化程度較高,比重大,顏色深,管胞較短。在橫切面上,管胞的孔徑多為圓形。伸張木,雙子葉植物的應力木。在傾斜或向上捲曲莖或枝條上側的木材,其特點是具膠質纖維,比正常木材木質化程度低,但纖維素含量較高。伸張木也存在於根中。
參考文獻
- ↑ 木材知識大全知乎
- ↑ 什麼是木質部?什麼是木質素?一米養殖